|
|
 |

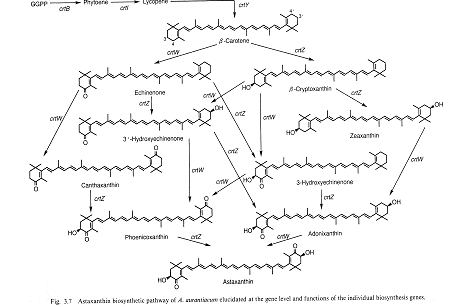
アスタキサンチンの細菌における生合成の経路は、Misawaを中心とした研究クループによる一連の研究の成果で解明された。β-カロテンからアスタキサンチンに至るカロテノイド類の生合成遺伝子を各種の微生物から遺伝子操作の手法を駆使して分離し、大腸菌系で発現解析することによりその生合成経路が明らかにされた。概要はMisawa 1) により次図にまとめられている(文献1より許可を得て引用)。
Misawaらの研究を中心に、一連の研究の主な文献を年代順に要約する。
H. pluvialisからβ-C-4 oxygenase (β-carotene ketolase)遺伝子をクローニングした。発現cDNAライブラリーをβ-カロテンを発現するように遺伝子操作したE. coliにtransfectし、目視て茶赤色な生産株を選別した。100,000コロニーからカンタキサンチンを作る色調が変化した1株を取得 2) 。この株はβ-カロテンからエキネノンをへてカンタキサンチンを作るβ-C-4 oxygenase活性を持っていた。この酵素はゼアキサンチンをアスタキサンチンへ変換しない。したがってH. pluvialisはカンタキサンチンをアスタキサンチンに変換する別の酵素を持つことが示唆された。
海性細菌Agrobacterium aurantiacumよりアスタキサンチンを生合成するカロテノイド合成遺伝子群を単離した。このクラスターはcrtW, crtZ, crtY, crtI, crtBの5コの関連遺伝子を含んでいた。crtBを除いて停止コドンは続く遺伝子の開始コドンとオバーラップしていた。カアロテノイド合成菌Erwinia uredovoraのカロテノイド生合成の基質を合成する遺伝子で形質転換したE. coliを用いてこれらの遺伝子の機能を検討し、アスタキサンチン生合成経路を提案した 3) 。β-カロテンからアスタキサンチンへの酸化反応を行うcrtW, crtZ遺伝子産物は、低い基質特異性を持っていた。したがって多くの中間体アドニキサンチン、フェニコキサンチン(アドニルビン)、カンタキサンチン、3'-ハイドロキシエキネノン、3-ハイドロキシエキネノンなども合成されていた。新たにH. pluvialisからアスタキサンチンを合成するcDNAを単離 4) し、それをE.uredovoraカロテノイド合成遺伝子で形質転換したE. coliで発現してアスタキサンチンを合成した。これらのクローンは、β-カロテンからエキネノンをへてカンタキサンチンへと変換する新規なβ-カロテンケトラーゼ(β-carotene oxygenase)をコードしていた。したがってこの酵素はメチレンをケト基に直接変換する酵素であった。mRNA分析からH. pluvialisでは cyst細胞のみで発現していた。同じくE.uredovora遺伝子で形質転換したゼアキサンチン合成能を持つE. coliを得られた新規cDNAでさらに形質転換した株でアスタキサンチン(3S, 3'S)が合成された。海洋性細菌Agrobacterium aurantiacumとAlcaligenes sp.strain PC-1のアスタキサンチン生合成遺伝子(crtZ と crtW)及びH. pluvialisのbktとE.uredovoraのcrtZ遺伝子を発現する形質転換E coliを用いて各酵素の性質を解析した 5) 。crtZ酵素は二つの酵素反応を触媒する、すなわちβ-カロテンからβ-クリプトキサンチンをへてゼアキサンチンの合成と、カンタキサンチンからフェニコキサンチン(アドニルビン)をへてアスタキサンチンの合成反応である。bkt,crtW酵素は、ゼアキサンチンから微量のアスタキサンチン生成を伴うが、β-カロテンと4-ケトゼアキサンチン(アドニキサンチン)からエキネノンをへてカンタキサンチンを合成する。海産細菌のcrtZ酵素は、カンタキサンチンをアスタキサンチンへ主に転換合成し、E.uredovoraのcrtZ酵素はβ-カロテンからゼアキサンチンを主に合成する。これらの酵素は酸素要求性である。活性に補助因子として2-oxoglutarate, ascorbic acid, Fe2+が関与する。解析の結果、crtZ-様酵素はcarotenoid 3, (3')- β-ionone ring hydroxylaseでcrtW/bkt酵素は carotenoid 4, (4')- β-ionone ring oxygenaseであった。また、E.uredovoraとA.aurantiacumからリコペン、β-カロテン、アスタキサンチンを生合成するcrt 遺伝子群を分離した 6) 。E.uredovoraのcrt がコードする酵素の基質は、カロテノイドやステロールの前駆体であるファルネシルピロ燐酸、ドリコール、他のイソプレノイド化合物などである。E coliをリコペン、β-カロテン、アスタキサンチンを蓄積するように形質転換できた。Zymomonas mobilisも形質転換できた。エンハンサー遺伝子もPh.rhodozymaやH.pluvialisから得られた。この遺伝子は、isopentenyl pyrophophate isomeraseをコードしていた。酵母由来のプロモーターとターミネーターを用いるとC.utilisやS.cerevisiaeでもカロテノイドを生合成できる。リコペンを生合成できるC.utilisやS.cerevisiae は、エルゴステロール含量が10-35%減少していた。代謝経路が変化したことを示唆している。C.utilisでは、HMGCoA還元酵素をコードする遺伝子の発現がカロテノイドレベルを増加させる。Arabidopsis thalianaのβ-carotene hydroxylase cDNAと相同なものをH.pluvialisからも分離された 7) 。推定アミノ酸配列は、 A.thalianaとC.annuumとは38%、原核生物とは32-34%の相同を示した。形質転換E. coliでH.pluvialisの酵素は、β-カロテンからゼアキサアンチン生成のみならずカンタキサンチンのアスタキサンチンへの転換も触媒した。このmRNAは、アスタキサンチン生合成誘導後上昇していることからH. pluvialisのcyst細胞でのアスタキサンチン合成に関与することを示唆する。
アスタキサンチンを、解明された酵素を用いて酵素的に合成することもできた 8) 。β-ヨノン環の3,3’-水酸化酵素(A.aurantiacumとAlcaligenes sp. PC-1)は、フェニコキサンチンをアスタキサンチンへ変換し、3-ハイドロキシエキネノンを4-ケトゼアキサンチンへ変換し、3’-ハイドロキシエキネノンを4-ケトゼアキサンチンへと変換した。同様に、3-および3’-ハイドロキシエキネノンをへてエキネノンを4-ケトゼアキサンチンへ変換した。4,4’-oxygenase(A. aurantiacum、Alcaligenes sp.PC-1、H.pluvialis)は、4-ケトゼアキサンチンをアスタキサンチンへ変換し、3-および3’-ハイドロキシエキネノンをフェニコキサンチンへ変換し、エキネノンをカンタキサンチンへと変換した。β-カロテンのβ-ヨノン環のモノ水酸化とケトン化によりアスタキサンチンが合成された。
遺伝子操作でタバコ(Nicotiana tabacum) でもアスタキサンチンが合成された 9) 。H.pluvialis crtO遺伝子(β-carotene hydroxylase)のcDNA を、トマトのphytoene desaturaseをプロモーターとしてタバコを形質転換した。 形質転換タバコは、(3S,3'S) アスタキサンチンと他のケトカロテノイドが蓄積し、黄色から赤に変色した。果実や花の色変換にも利用できる可能性を示唆される。

アスタキサンチン生合性に関する文献
|
|